![]()
Reference to Wikipedia (Taken From http://en.wikipedia.org/wiki/Brodmann_area_41_%26_42 )
Brodmann area 41
This area is also known as anterior transverse temporal area 41 (H). It is a subdivision of the cytoarchitecturally-defined temporal region of cerebral cortex, occupying the anterior transverse temporal gyrus (H) in the bank of the lateral sulcus on the dorsal surface of the temporal lobe. Brodmann area 41 is bounded medially by the parainsular area 52 (H) and laterally by the posterior transverse temporal area 42 (H) (Brodmann-1909).
Brodmann area 42
This area is also known as posterior transverse temporal area 42 (H). It is a subdivision of the cytoarchitecturally-defined temporal region of cerebral cortex, located in the bank of the lateral sulcus on the dorsal surface of the temporal lobe. Brodmann area 42 is bounded medially by the anterior transverse temporal area 41 (H) and laterally by the superior temporal area 22 (Brodmann-1909).
Relationship to auditory system

The auditory cortex is the most highly organized processing unit of sound in the brain. This cortex area is the neural crux of hearing, and, in humans, language and music.
The auditory cortex is divided into three separate parts, the primary, secondary and tertiary auditory cortex. These structures are formed concentrically around one another, with the primary AC in the middle and the tertiary AC on the outside.
The primary auditory cortex is tonotopically organized, which means that certain cells in the auditory cortex are sensitive to specific frequencies. This is a fascinating function which has been preserved throughout most of the audition circuit. This area of the brain "is thought to identify the fundamental elements of music, such as pitch and loudness." [4] This makes sense as this is the area which receives direct input from the medial geniculate nucleus of the thalamus. The secondary auditory cortex has been indicated in the processing of "harmonic, melodic and rhythmic patterns." [4] The tertiary auditory cortex supposedly integrates everything into the overall experience of music. [4]
An evoked response study of congenitally deaf kittens by Klinke et al. utilized field potentials to measure cortical plasticity in the auditory cortex. These kittens were stimulated and measured against a control or un-stimulated congenitally deaf cat (CDC) and normal hearing cats. The field potentials measured for artificially stimulated CDC was eventually much stronger than that of a normal hearing cat. [5] This is in concordance with Eckart Altenmuller's study where it was observed that students who received musical instruction had greater cortical activation than those who did not. [6]
The auditory cortex exhibits some strange behavior pertaining to the gamma wave frequency. When subjects are exposed to three or four cycles of a 40 hertz click, an abnormal spike appears in the EEG data, which is not present for other stimuli. The spike in neuronal activity correlating to this frequency is not restrained to the tonotopic organization of the auditory cortex. It has been theorized that this is a " resonant frequency" of certain areas of the brain, and appears to affect the visual cortex as well. [7]
Gamma band activation (20 to 40 Hz) has been shown to be present during the perception of sensory events and the process of recognition. Kneif et al, in their 2000 study, presented subjects with eight musical notes to well known tunes, such as Yankee Doodle and Frere Jacques. Randomly, the sixth and seventh notes were omitted and an electroencephalogram, as well as a magnetoencephalogram were each employed to measure the neural results. Specifically, the presence of gamma waves, induced by the auditory task at hand, were measured from the temples of the subjects. The OSP response, or omitted stimulus response, was located in a slightly different position; 7 mm more anterior, 13 mm more medial and 13 mm more superior in respect to the complete sets. The OSP recordings were also characteristically lower in gamma waves, as compared to the complete musical set. The evoked responses during the sixth and seventh omitted notes are assumed to be imagined, and were characteristically different, especially in the right hemisphere. [8] The right auditory cortex has long been shown to be more sensitive to tonality, while the left auditory cortex has been shown to be more sensitive to minute sequential differences in sound specifically speech.
Hallucinations have been shown to produce oscillations which are parallel (although not exactly the same as) the gamma frequency range. Sperling showed in his 2004 study that auditory hallucinations produce band wavelengths in the range of 12.5-30 Hz. The bands occurred in the left auditory cortex of a schizophrenic and were controlled against 13 controls (18) . This aligns with the studies of people remembering a song in their minds; they do not perceive any sound, but experience the melody, rhythm and overall experience of sound. When schizophrenics experience hallucinations, it is the primary auditory cortex which becomes active. This is characteristically different from remembering a sound stimulus, which only faintly activates the tertiary auditory cortex. [9] By deduction, an artificial stimulation of the primary auditory cortex should elicit an incredibly real auditory hallucination. The termination of all audition and music into the tertiary auditory cortex creates a fascinating nexus of aural information. If this theory is true, it would be interesting to study a subject with a damaged, TAC or one with artificially suppressed function. This would be very difficult to do as the tertiary cortex is simply a ring around the secondary, which is a ring around the primary AC.
Tone is perceived in more places than just the auditory cortex; one specifically fascinating area is the rostromedial prefrontal cortex.[10] Janata et al, in their 2002 study, used an fMRI machine to study the areas of the brain which were active during tonality processing. The result of which displayed several areas which are not normally considered to be part of the audition process. The rostromedial prefrontal cortex is a subsection of the medial prefrontal cortex, which projects to the amygdala , and is thought to aid in the inhibition of negative emotion. [11] The medial prefrontal cortex is thought to be the core developmental difference between the impulsive teenager and the calm adult. The rostromedial prefrontal cortex is tonality sensitive, meaning it is activated by the tones and frequencies of resonant sounds and music. It could be hypothesized that this is the mechanism by which music ameliorates the soul (or, if one prefers, the limbic system).1 - intermediate postcentral (area postcentralis intermedia). Located in the postcentral gyrus. Bounded cytoarchitecturally by the rostral postcentral area 3 and the caudal postcentral area 2 and, at its ventral tip, by the subcentral area 43.
2 - caudal postcentral (area postcentralis caudalis). Located primarily in the caudal portion of the postcentral gyrus and the rostral lip of the postcentral sulcus with a caudal extension along the intraparietal sulcus. Cytoarchitecturally bounded rostrally by the intermediate postcentral area 1 and caudally by the preparietal area 5, the superior parietal area 7 and the supramarginal area 40.
Figure 1. Courtesy of Brodmann Brain Picture from Ruye Wang 1999-08-31

5 - preparietal (area praeparietalis). Occupies the superior parietal lobule and a portion of the postcentral gyrus, particularly on the medial aspect of the hemisphere. Bounded approximately by the cingulate sulcus on the medial aspect of the hemisphere and by the superior postcentral sulcus on the lateral aspect. Cytoarchitecturally bounded by the caudal postcentral area 2, the superior parietal area 7 and on the medial bank of the hemisphere by the gigantopyramidal area 4 and the dorsal posterior cingulate area 31.
6 - agranular frontal (area frontalis agranularis). Located primarily in the caudal portions of the superior frontal gyrus and the middle frontal gyrus and the rostral portions of the precentral gyrus not occupied by the gigantopyramidal area 4. It extends from the cingulate sulcus on the medial aspect of the hemisphere to the lateral sulcus on the lateral aspect. Cytoarchitecturally bounded rostrally by the frontal region and caudally by the gigantopyramidal area 4.
7 - superior parietal (area parietalis superior). Occupies much of the superior parietal lobule and some of the precuneus. Bounded approximately by the superior postcentral sulcus rostrally, the intraparietal sulcus laterally, the parieto-occipital sulcus caudally and, on the medial bank of the hemisphere, the subparietal sulcus. Cytoarchitecturally bounded rostrally by the preparietal area 5 and the caudal postcentral area 2; caudally by the peristriate area 19; and medially by the dorsal posterior cingulate area 31.
8 - intermediate frontal (area frontalis intermedia). Located primarily in the superior frontal gyrus extending from the cingulate sulcus on the medial surface over the margin of the hemisphere to the middle frontal gyrus. Cytoarchitecturally bounded caudally by the agranular frontal area 6 and ventrally by the granular frontal area 9.
9 - granular frontal (area frontalis granularis). Occupies portions of the superior frontal gyrus and the middle frontal gyrus. Its approximate boundary on the medial aspect of the hemisphere is the cingulate sulcus and, on the lateral aspect, the inferior frontal sulcus. Cytoarchitecturally bounded dorsocaudally by the intermediate frontal area 8, caudally by the agranular frontal area 6, and ventrally by the frontopolar area 10, the middle frontal area 46 and the opercular area 44.
10 - frontopolar (area frontopolaris). Occupies the most rostral portions of the superior frontal gyrus and the middle frontal gyrus. On the medial aspect of the hemisphere it is bounded ventrally by the superior rostral sulcus. It does not extend as far as the cingulate sulcus. Cytoarchitecturally bounded dorsally by the granular frontal area 9, caudally by the middle frontal area 46, and ventrally by the orbital area 47 and by the frontopolar area 12.
11 - prefrontal (area praefrontalis). Constitutes most of the orbital gyri, gyrus rectus and the most rostral portion of the superior frontal gyrus. Bounded medially by the inferior rostral sulcus and laterally approximately by the frontomarginal sulcus. Cytoarchitecturally bounded on the rostral and lateral aspects of the hemisphere by the frontopolar area 10, the orbital area 47, and the triangular area 45; on the medial surface it is bounded dorsally by the area 12 and caudally by the subgenual area 25.
12 - prefrontal (area praefrontalis). Occupies the area between the superior rostral sulcus and the inferior rostral sulcus. Cytoarchitecturally bounded dorsally by the frontopolar area 10 and the dorsal anterior cingulate area 32; caudally, ventrally and rostrally it is bounded by the prefrontal area 11. (Originally described as part of prefrontal area 11 but not shown in the map, subsequently it was labeled as an independent area 12.)
17 - striate (area striata). Part of the occipital lobe of the cerebral cortex that is defined on the basis of cyto- and myeloarchitecture, primarily by the band/stripe of Gennari.
18 - parastriate (area parastriata). Located in parts of the cuneus, the lingual gyrus and the lateral occipital gyrus of the occipital lobe. Cytoarchitecturally bounded on one side by the striate area 17, from which it is distinguished by absence of a band/stripe of Gennari, and on the other by the peristriate area 19.
19 - peristriate (area peristriata). Located in parts of the lingual gyrus, the cuneus, the lateral occipital gyrus and the superior occipital gyrus of the occipital lobe where it is bounded approximately by the parieto-occipital sulcus. Cytoarchitecturally bounded on one side by the parastriate area 18 which it surrounds. Rostrally it is bounded by the angular area 39 and the occipitotemporal area 37.
20 - inferior temporal (area temporalis inferior). Corresponds approximately to the inferior temporal gyrus. Cytoarchitecturally bounded medially by the ectorhinal area 36, laterally by the middle temporal area 21, rostrally by the temporopolar area 38 and caudally by the occipitotemporal area 37.
21 - middle temporal (area temporalis media). Corresponds approximately to the middle temporal gyrus. Bounded rostrally by the temporopolar area 38, ventrally by the inferior temporal area 20, caudally by the occipitotemporal area 37, and dorsally by the superior temporal area 22.
22 - superior temporal (area temporalis superior). Corresponds approximately to the lateral and caudal two thirds of the superior temporal gyrus. Bounded rostrally by the temporopolar area 38, medially by the posterior transverse temporal area 42, ventrocaudally by the middle temporal area 21 and dorsocaudally by the supramarginal area 39.
23 - ventral posterior cingulate (area cingularis posterior ventralis). Occupies most of the posterior cingulate gyrus adjacent to the corpus callosum. At the caudal extreme it is bounded approximately by the parieto-occipital sulcus. Cytoarchitecturally bounded dorsally by the dorsal posterior cingulate area 31, rostrally by the ventral anterior cingulate area 24, and ventrorostrally in its caudal half by the retrosplenial region.
24 - ventral anterior cingulate (area cingularis anterior ventralis). Occupies most of the anterior cingulate gyrus in an arc around the genu of corpus callosum. Its outer border corresponds approximately to the cingulate sulcus. Cytoarchitecturally bounded internally by the pregenual area 33, externally by the dorsal anterior cingulate area 32, and caudally by the ventral posterior cingulate area 23 and the dorsal posterior cingulate area 31.
25 - subgenual (area subgenualis). A narrow band located in the caudal portion of the subcallosal area adjacent to the paraterminal gyrus from which it is separated by the posterior parolfactory sulcus. It is bounded by the prefrontal area 11rostrally and by the paraterminal gyrus caudally.
26 - ectosplenial (area ectosplenialis). A narrow band located in the isthmus of cingulate gyrus adjacent to the fasciolar gyrus internally. It is bounded externally by the granular retrolimbic area 29.
28 - entorhinal (area entorhinalis). Located in the entorhinal area on the medial aspect of the temporal lobe. It and the dorsal entorhinal area 34 together constitute approximately the entorhinal area.
29 - granular retrolimbic (area retrolimbica granularis). A narrow band located in the isthmus of cingulate gyrus. Cytoarchitecturally bounded internally by the ectosplenial area 26 and externally by the agranular retrolimbic area 30.
30 - agranular retrolimbic (area retrolimbica agranularis). Located in the isthmus of cingulate gyrus. Cytoarchitecturally bounded internally by the granular retrolimbic area 29, dorsally by the ventral posterior cingulate area 23 and ventrolaterally by the ectorhinal area 36.
31 - dorsal posterior cingulate (area cingularis posterior dorsalis). Occupies portions of the posterior cingulate gyrus and medial aspect of the parietal lobe. Approximate boundaries are the cingulate sulcus dorsally and the parieto-occipital sulcus caudally. It partially surrounds the subparietal sulcus. Cytoarchitecturally bounded rostrally by the ventral anterior cingulate area 24, ventrally by the ventral posterior cingulate area 23, dorsally by the gigantopyramidal area 4 and preparietal area 5 and caudally by the superior parietal area 7.
32 - dorsal anterior cingulate (area cingularis anterior dorsalis). Forms an outer arc around the anterior cingulate gyrus. The cingulate sulcus defines approximately its inner boundary and the superior rostral sulcus its ventral boundary; rostrally it extends almost to the margin of the frontal lobe. Cytoarchitecturally bounded internally by the ventral anterior cingulate area 24, externally by medial margins of the agranular frontal area 6, intermediate frontal area 8, granular frontal area 9, frontopolar area 10, and prefrontal area 11.
33 - pregenual (area praegenualis). A narrow band located in the anterior cingulate gyrus adjacent to the supracallosal gyrus in the depth of the callosal sulcus. Cytoarchitecturally bounded by the ventral anterior cingulate area 24 and the supracallosal gyrus.
34 - dorsal entorhinal (area entorhinalis dorsalis). Located in the entorhinal area on the medial aspect of the temporal lobe. It and the entorhinal area 28 together constitute approximately the entorhinal area.
35 - perirhinal (area perirhinalis). Located along the rhinal sulcus. Cytoarchitectually bounded medially by the entorhinal area 28 and laterally by the ectorhinal area 36.
36 - ectorhinal (area ectorhinalis). Located primarily in the fusiform gyrus, with its medial boundary corresponding approximately to the rhinal sulcus. Cytoarchitecturally bounded laterally and caudally by the inferior temporal area 20, medially by the perirhinal area 35 and rostrally by the temporopolar area 38.
37 - occipitotemporal (area occipitotemporalis). Located primarily in the caudal portions of the fusiform gyrus and inferior temporal gyrus on the mediobasal and lateral surfaces at the caudal extreme of the temporal lobe. Cytoarchitecturally bounded caudally by the peristriate area 19, rostrally by the inferior temporal area 20 and middle temporal area 21 and dorsally on the lateral aspect of the hemisphere by the angular area 39.
38 - temporopolar (area temporopolaris). Located primarily in the most rostral portions of the superior temporal gyrus and the middle temporal gyrus. Cytoarchitecturally bounded caudally by the inferior temporal area 20, the middle temporal area 21, the superior temporal area 22 and the ectorhinal area 36.
39 - angular (area angularis). Corresponds to the angular gyrus surrounding the caudal tip of the superior temporal sulcus. Dorsally it is bounded approximately by the intraparietal sulcus. Cytoarchitecturally bounded rostrally by the supramarginal area 40, dorsally and caudally by the peristriate area 19, and ventrally by the occipitotemporal area 37.
40 - supramarginal (area supramarginalis). Located primarily in the supramarginal gyrus surrounding the posterior ascending limb of lateral sulcus. Bounded approximately by the intraparietal sulcus, the inferior postcentral sulcus the posterior subcentral sulcus and the lateral sulcus. Cytoarchitecturally bounded caudally by the angular area 39, rostrally and dorsally by the caudal postcentral area 2, and ventrally by the subcentral area 43 and the superior temporal area 22.
41 - anterior transverse temporal (area temporalis transversa anterior). Occupies the anterior transverse temporal gyrus in the bank of the lateral sulcus on the dorsal surface of the temporal lobe. Cytoarchitecturally bounded medially by the parainsular area 52 and laterally by the posterior transverse temporal area 42.
42 - posterior transverse temporal (area temporalis transversa posterior). Located in the bank of the lateral sulcus on the dorsal surface of the temporal lobe. Cytoarchitecturally bounded medially by the anterior transverse temporal area 41 and laterally by the superior temporal area 22.
43 - subcentral (area subcentralis). Occupies the postcentral gyrus and the precentral gyrus between the ventrolateral extreme of the central sulcus and the depth of the lateral sulcus at the insula. Its rostral and caudal borders are approximated by the anterior subcentral sulcus and the posterior subcentral sulcus respectively. Cytoarchitecturally bounded rostrally by the agranular frontal area 6 and caudally, for the most part, by the caudal postcentral area 2 and the supramarginal area 40.
44 - opercular (area opercularis). Corresponds approximately to the opercular part of the inferior frontal gyrus. Bounded caudally by the inferior precentral sulcus and rostrally by the anterior ascending limb of lateral sulcus. It surrounds the diagonal sulcus. In the depth of the lateral sulcus it borders on the insula. Cytoarchitectonically bounded caudally and dorsally by the agranular frontal area 6, dorsally by the granular frontal area 9 and rostrally by the triangular area 45.
45 - triangular (area triangularis). Occupies the triangular part of the inferior frontal gyrus and, surrounding the anterior horizontal limb of lateral sulcus, a portion of the orbital part of inferior frontal gyrus. Bounded caudally by the anterior ascending limb of lateral sulcus, it borders on the insula in the depth of the lateral sulcus . Cytoarchitectonically bounded caudally by the opercular area 44, rostrodorsally by the middle frontal area 46 and ventrally by the orbital area 47.
46 - middle frontal (area frontalis media). Occupies approximately the middle third of the middle frontal gyrus and the most rostral portion of the inferior frontal gyrus. Cytoarchitecturally bounded dorsally by the granular frontal area 9, rostroventrally by the frontopolar area 10 and caudally by the triangular area 45.
47 - orbital (area orbitalis). Surrounds the caudal portion of the orbital sulcus from which it extends laterally into the orbital part of inferior frontal gyrus. Cytoarchitectonically bounded caudally by the triangular area 45, medially by the prefrontal area 11, and rostrally by the area frontopolaris 10.
48 - retrosubicular (area retrosubicularis). Located on the medial surface of the temporal lobe. Cytoarchitectually bounded rostrally by the perirhinal area 35 and medially by the presubiculum. (While described by Brodmann, it was not included in his areal maps of human cortex.)
52 - parainsular (area parainsularis) Located in the bank of the lateral sulcus on the dorsal surface of the temporal lobe. Its medial boundary corresponds approximately to the junction between the temporal lobe and the insula. Cytoarchitecturally bounded laterally by the anterior transverse temporal area 41.
13, 14, 15, 16, 27, 49, 50, 51 - monkey only

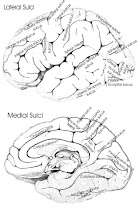
Taken from Prof Mark Dubin of University of Colorado

Taken from
Professor Mark Dubin of University of Colorado on Functional attribution of Brodmann Areas, with special attention to language processing
Lables of each area, see picture below. Picture is taken from
Professor Mark Dubin of University of Colorado 
Reference to Wikipedia
Brodmann areas were originally defined by Korbinian Brodmann and referred to by numbers from 1 to 52. It is obtained by the organization of the cortex as observed when a tissue is stained for nerve cells called cytoarchitecture.
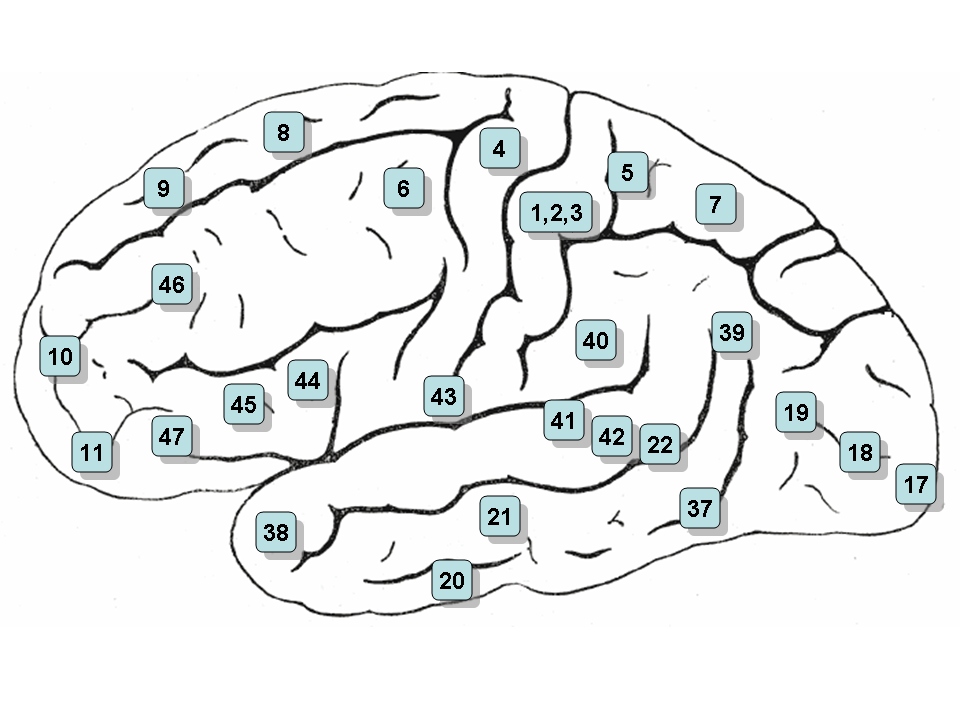 Areas 1, 2 & 3 - Primary Somatosensory Cortex
Areas 1, 2 & 3 - Primary Somatosensory Cortex
Area 5 - Somatosensory Association Cortex
Area 6 - Pre-Motor and Supplementary Motor Cortex (Secondary Motor Cortex)
Area 7 - Somatosensory Association Cortex
Area 8 - Includes Frontal eye fields
Area 9 - Dorsolateral prefrontal cortex
Area 10 - Frontopolar area (most rostral part of superior and middle frontal gyri)

Area 11 - Orbitofrontal area (orbital and rectus gyri, plus part of the rostral part of the superior frontal gyrus)
Area 12 - Orbitofrontal area (used to be part of BA11, refers to the area between the superior frontal gyrus and the inferior rostral sulcus)
Area 13 and Area 14* - Insular cortex
Area 15* - Anterior Temporal Lobe
Area 17 - Primary Visual Cortex (V1)
Area 18 - Visual Association Cortex (V2)
Area 19 - V3
Area 20 - Inferior Temporal gyrus
Area 21 - Middle Temporal gyrus
Area 22 - Superior Temporal Gyrus, of which the rostral part participates to Wernicke's area
Area 23 - Ventral Posterior cingulate cortex
Area 24 - Ventral Anterior cingulate cortex
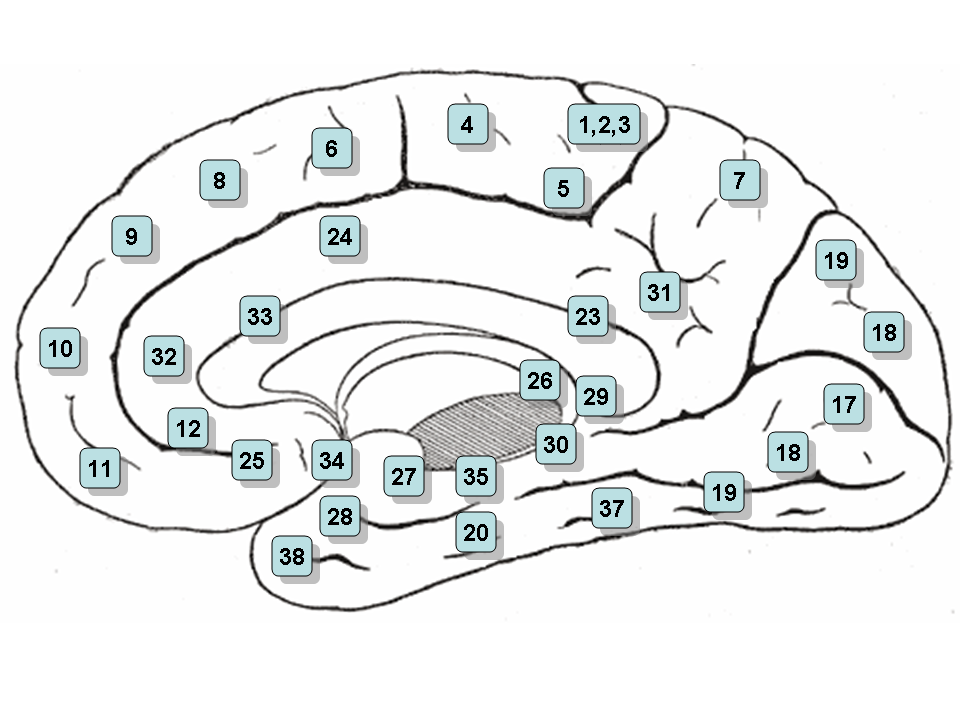 Area 25 - Subgenual cortex
Area 25 - Subgenual cortexArea 26 - Ectosplenial area
Area 28 - Posterior Entorhinal Cortex
Area 29 - Retrosplenial cingular cortex
Area 30 - Part of cingular cortex
Area 31 - Dorsal Posterior cingular cortex
Area 32 - Dorsal anterior cingulate cortex
Area 34 - Anterior Entorhinal Cortex (on the Parahippocampal gyrus)
Area 35 - Perirhinal cortex (on the Parahippocampal gyrus)
Area 36 - Parahippocampal cortex (on the Parahippocampal gyrus)
Area 37 - Fusiform gyrus
Area 38 - Temporopolar area (most rostral part of the superior and middle temporal gyri
Area 39 - Angular gyrus, part of Wernicke's area
Area 40 - Supramarginal gyrus part of Wernicke's area
Areas 41 & 42 - Primary and Auditory Association Cortex
Area 43 - Subcentral area (between insula and post/precentral gyrus)
Area 44 - pars opercularis, part of Broca's area
Area 45 - pars triangularis Broca's area
Area 46 - Dorsolateral prefrontal cortex
Area 47 - Inferior prefrontal gyrus
Area 48 - Retrosubicular area (a small part of the medial surface of the temporal lobe)
Area 52 - Parainsular area (at the junction of the temporal lobe and the insula)
From Wikipedia, http://en.wikipedia.org/wiki/Brodmann_area Fig 1 & 3 are faithful reproduction of a lithograph plate from Gray's Anatomy

Functional areas for face recognition and face processing.
Figure 2: Brain regions central in face perception, as outlined and studied in this thesis.
Upper left: typical MRI-image of the right side of the brain, displaying several brain regions important in face perception. Locations of the coronal sections in A, B and C are shown.
Amy=amygdala; IFG=inferior frontal gyrus; Ins=insula; latFG=lateral fusiform
gyrus; pIPG=posterior inferior parietal gyrus; STS=superior temporal sulcus.
 Several brain regions central infacial expression production, as outlinedand studied in this thesis. Left: typical MRIimageof the right side of the brain. Right:typical MRI image of the brain from above.M1=primary motorcortex; PM=premotorcortex; SMA=supplementary motor area;pre-SMA=pre-supplementary motor area.
Several brain regions central infacial expression production, as outlinedand studied in this thesis. Left: typical MRIimageof the right side of the brain. Right:typical MRI image of the brain from above.M1=primary motorcortex; PM=premotorcortex; SMA=supplementary motor area;pre-SMA=pre-supplementary motor area.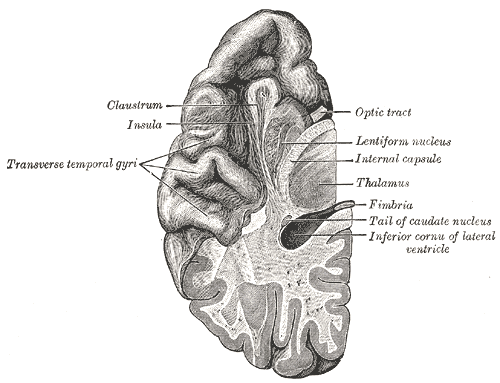
Finally, I have got it submitted after re-printing and re-binding the whole thesis.
Because, Pak said I should use font size 12 instead of 10. And because I had a messy reference, missing this and that and in new and old APA format...
And my figures were all in black & white.
So... when the Faculty refused to take my thesis because I have not got all my 4 supervisors' signatures to support my submission... I was initially very frustrated.. and then I ...
Get this chance to re-do the reference, font size, color figures, and RE-print.. What a waste of paper and trees!
I should have handed in at 5pm, but I only managed to finish binding it at 6.45pm... it was all very hectic!
The themo-binding maching needs 30 MINS to warm UP... oh dear... so I stood there waiting waiting nervously.........
and the Faculty kept ringing me up to submit my thesis...or else I'd be disqualified...
I still have a lot of missing reference.....................ai ai ai!!!!!!!!!!!!!!!!!!!!!!!!!11
Connectivity hypothsis
Connectivity has been implicated as a central abnormality in schizophrenia.
Definitive pathophysiologica basis for abnromal connectivity remeains elusive.
Only until recently whit matter, ie oligodendroglia and myelin, in the pathogenesis of schizophrenia. The cellular and molecular abnormalities found in Sz brain.
Neuronal connectivity affected profoundly by
- myelination
- factors that affect myelination (oligodendroglia function)
- quality of myelin deposition (affected by oligodendroglia function)
- synaptic transmission (also affected by oligodendroglia function)
Davis et al. 2003 White Matter Changes in Sz. Evidence for Myelin-related dysfunction. Arch Gen Psyc 2003:60:443-456.








The dopamine pathways
The dopamine hypothesis of schizophrenia proposed that schizophrenia is associated with an increase in dopamine transmission between the substantia nigra to the caudate nucleus-putamen (neostriatum) compared with normal, while in the other major dopaminergic pathways to the mesolimbic forebrain and the tubero-infundibular system dopamine transmission is reduced.
[The nigrostriatal and mesolimbic dopamine tracts. In: Basic concepts in Neuroscience, international edition. Slaughter M. London, McGraw-Hill Medical Publishing Division 2002,187–208 ]
The glutamate pathways
Hypoactive glutamate pathways are believed to be associated with psychosis and confusion in schizophrenia. These pathways include: cortico-cortical pathways, thalamic-cortical, extrapyramidal pathways (cortical-striatum), and projections exist between the cortex, substantia nigra, subthalmic nucleus and pallidum.
[Brain structures and neuronal projections. Institute of Chemistry, Pharmacology and Biomedical Sciences. University of Sunderland. http://abdellab.sunderland.ac.uk/lectures/Brain/glu1.html. Accessed on 21 March 2003]
The serotonergic pathway
Up-regulation of the two serotonergic pathways: the dorsal raphe nuclei project into the substantia nigra and
the projections from the rostral raphe nuclei ascending into the cerebral cortex, limbic regions and basal ganglia
--> leads to hypofunction of the dopaminergic system, and this effect may be responsible for the negative symptoms of schizophrenia.
[The role or serotonin in schizophrenia. The American college of neuropsychopharmacology. www.acnp.org/g4/GN401000117/CH115.html. Accessed on 21 March 2003. ]
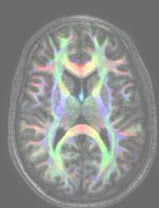
Schizophrenia is a major mental illness thought to result from disordered neurodevelopment. It is postulated that connectivity within brain systems controlling perception, language and cognition is disrupted. Using Diffusion Tensor Imaging (DTI) the integrity of white matter pathways in brain can be assessed. The directional diffusion of water molecules (fractional anisotrophy, FA) ranges from 0 (completely random) to 1 (completely isotrophic in one direction) such that the more constrained movement is, say within a myelin sheath, the closer fractional anisotrophy is to 1. However, DTI studies in schizophrenia are confounded by medication effects and the duration of illness. To avoid these complications this study is one of the first to investigate FA indices of cerebral white matter connectivity in never-medicated individuals with first onset schizophrenia. White matter abnormalities in patients with schizophrenia were first compared to matched healthy subjects, and then correlations with illness severity examined. Twenty-five never-medicated patients with first episode schizophrenia and 24 healthy volunteers completed the study. The two groups did not significantly differ in age, sex ratio, handedness or ethnicity. Voxel-based analysis (VBA) was used to compare fractional anisotropy (FA) maps between the schizophrenia group and healthy controls. The VBA analyses were repeated with various sizes of smoothing kernel (4mm to 14mm) and with both parametric and non-parametric statistical approaches, in order to determine the most reliable result. A region of interest (ROI) analysis of FA scores was performed on 6 brain regions related to the significant group difference found in VBA, blind to group membership, to further assess validity of parametric and non-parametric methods. Results obtained consistently showed FA in the patients was not increased relative to healthy controls, but reduced in the right posterior limb of internal capsule, left middle temporal gyrus, right substantia nigra, left cerebral peduncle, and splenium of corpus callosum. Voxel-wise correlation analysis of the severity of positive and negative symptoms on the Positive and Negative Symptom Scale (PANSS) and FA in the patient group was performed. Selection of the most appropriate level of smoothing was based upon parametric VBA results from the group analysis. The results showed a significant positive correlation between positive symptom scores and FA values in bilateral medial frontal gyrus (left superior & right anterior region of coronal radiata), left cuneus (left posterior region of coronal radiata), left pre-central gyrus (superior occipitofrontal fasciculus), right body of cingculum, right medial orbital gyrus (inferior frontooccipital fasciculus/uncinate fasciculus), left superior longitudinal fasciculus near insular gyrus, and left superior temporal gyrus (inferior longitudinal fasciculs/inferior frontooccipital fasciculus). In conclusion, this is the first study to demonstrate that striato-limbic white matter dysconnectivity in schizophrenia is unrelated to treatment and illness chronicity. The use of FA as a useful marker for pathology in schizophrenia as well as its potential to assess response to neuroleptic treatment is discussed.
![]()


{kind=link}